April 25, 2005
News about brain structure in Williams Syndrome
 In the latest Journal of Neuroscience, there's an interesting paper about
brain structure in Williams Syndrome, a disorder caused by deletions of variable length in a gene on chromosome 7 (7q11.23) that codes for the connective-tissue protein elastin, and perhaps in other
adjacent genes. Among the many symptoms of the syndrome are mental redardation with hypersociability, relatively spared language, and relatively spared musical abilities that sometimes rise to savant levels. What's new in this paper is a systematic and thoughtful examination of differences in brain structure between WS subjects and controls.
In the latest Journal of Neuroscience, there's an interesting paper about
brain structure in Williams Syndrome, a disorder caused by deletions of variable length in a gene on chromosome 7 (7q11.23) that codes for the connective-tissue protein elastin, and perhaps in other
adjacent genes. Among the many symptoms of the syndrome are mental redardation with hypersociability, relatively spared language, and relatively spared musical abilities that sometimes rise to savant levels. What's new in this paper is a systematic and thoughtful examination of differences in brain structure between WS subjects and controls.
The reference is Thompson PM, Lee AD, Dutton RA, Geaga JA, Hayashi KM, Eckert MA, Bellugi U, Galaburda AM, Korenberg JR, Mills DL, Toga AW, Reiss AL. "Abnormal Cortical Complexity and Thickness Profiles Mapped in Williams Syndrome." Journal of Neuroscience, 25(16):4146-4158, April 20, 2005.
The background finding (in keeping with earlier studies) is one of general reduction in brain size, and especially in "white matter" (i.e. neuronal interconnections consisting of myelinated nerve fibers, as opposed to "grey matter", consisting mainly of cell bodies):
...the WS group had greatly reduced overall brain volumes (...left and right hemisphere volumes were reduced by 13.3 and 12.2%, respectively)...
... this overall deficit was attributable to primarily a far more dramatic reduction in white matter (left hemisphere, -18.0%...; right hemisphere, -18.3% ...) than gray matter, although gray matter also was reduced severely (left hemisphere, -6.8%...; right hemisphere, -6.2%...).
[The] WM deficit was found somewhat uniformly across all lobes (frontal, -15.9%; parietal, -18.4%; temporal, -21.2%; occipital, -20.0%...). Lobar GM volumes also appeared uniformly reduced (frontal, -5.9%; parietal, -7.4%; temporal, -5.3%; occipital, -8.6%).
However, against this background, there were striking local exceptions:
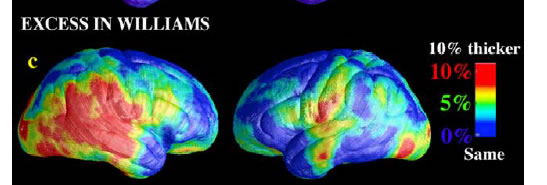
The WS group had greatly increased cortical thickness in a large neuroanatomical region encompassing the perisylvian language-related cortex. This region surrounds the posterior limit of the Sylvian fissures and extends inferiorly into the lateral temporal lobes (Figure 4c, red colors denote a 10% thickening of the cortex relative to controls). The region of significant thickness increases also extended over the inferior surface of the right temporal lobe (Fig. 4e) into the collateral and entorhinal cortex. This region included the fusiform face area, which processes facial stimuli, a cognitive ability in which WS subjects show notable strengths.
Here's the picture from their Figure 4c:
The authors also
...developed an algorithm to measure the fractal dimension, or complexity, of the human cerebral cortex, based on a previous algorithm that we developed for mapping the complexity of deep sulcal surfaces in the brain (Thompson et al., 1996).
According to this measure,
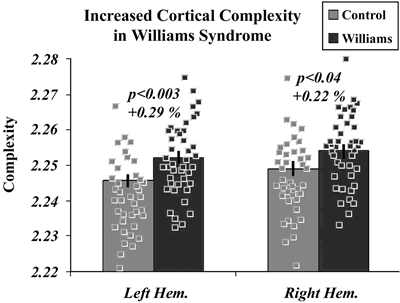
Cortical complexity was ... significantly increased in WS for both brain hemispheres.
The differences were small (e.g. left hemisphere Williams 2.2522 +/- 0.0016 SE, left hemisphere controls 2.2457 +/- 0.0014) but significant (p < 0.00145; two-tailed t test in this case). The authors comment that
Although these differences appear to be small in magnitude (~0.1– 0.3%), this can be misleading because they are computed from a log–log plot, in which small differences in slope translate into very large differences in gyral complexity.
As their plot shows, there is a great deal of overlap in the values for individual subjects:
Their discussion of function interpretations is interesting:
One simplistic interpretation is that thicker cortex is better, and that regionally thicker language cortex in WS subjects may account for their verbal strengths and unusually expressive language. By a similar argument, WS subjects are also prone to seek the gaze of others (Mervis et al., 2003), and the thicker cortical region in WS also encompasses the superior temporal sulcus, an area important in face and gaze processing (Kanwisher et al., 1997; Zeineh et al., 2003). However, this interpretation is unduly simplistic for several reasons. First, WS subjects have relatively intact language, but they do not outperform controls, which would be implied by the idea that thicker cortex is better (Haier et al., 2004). Second, WS subjects do not have enhanced function in other systems with thicker cortex (e.g., posterior and lateral occipital and inferior occipital-temporal regions), which subserve visuospatial functions impaired in WS. Third, a similar thickening of perisylvian cortex in fetal alcohol syndrome (FAS) (Sowell et al., 2002b) is not associated with better language function.
They add that
In both WS and FAS, excess cortical gray matter is most likely a result of a failure of cortical formation during gyrogenesis or a concomitant failure or delay in myelination, perhaps specifically in subcortical U-fibers (conventional MRI cannot distinguish these two possibilities).
In thinking about genetically-related brain structure/function correlations, I always wonder how much of such patterns is genetically determined in an open-loop sort of way, and how much depends on developmental processes that involve feedback from experience (which might amplify initial differences in ability, interest and motivation). So I was very happy to see the authors explicitly raise those issues:
Cortical architecture in WS is likely influenced by haploinsufficiency for specific deleted genes, but it is also dynamic and environmentally influenced throughout life. This study correlates genetic mutation and anatomical change, but causality cannot be determined. There is no way to make any simple categorical interpretation of genetic and nongenetic influences, because these cannot be disentangled, and both may occur downstream of a genetic lesion. The observed cortical thinning may be shaped primarily by negative genetic influences (that impair parietaloccipital structure and function). Nonetheless, the cortical increases may represent increased use or overuse of specific networks. Even if the thickening represents an adaptive response to the genetic deletion, whether or not it is functionally advantageous cannot be assessed without additional testing. Conversely, a cortex that appears relatively intact on MRI is not necessarily functionally intact. The notion that there are functions left intact in developmental disorders is likely incorrect; massive reorganization is likely standard across developmental disorders, and the resultant functionality is probably deviant (Karmiloff-Smith et al., 1997; Mills et al., 2000; Thomas and Karmiloff-Smith, 2002; Grice et al., 2003). In particular, language processing, musical abilities, and face processing in WS are not par with normal performance (Karmiloff-Smith et al., 1998, 2004).
Overall, this is fascinating work, not least because it's a welcome corrective to the simplistic interepretations that are sometimes given to the relative sparing of linguistic abilities in this syndrome.
Posted by Mark Liberman at April 25, 2005 11:51 AM